| |
|
|
| |
|
|
| Thèse |
|

Vive le travail de terrain!
|
J'ai effectué ma thèse sous la
direction de Jean-Claude Menaut à l'Ecole Normale Supérieure
dans un laboratoire d'écologie commun à l'ENS et à
Paris VI (à l'époque UMR 7625). Ce travail a été
réalisé en étroite collaboration avec Jacques
Gignoux qui m'a initié à l'écologie spatiale
et à la recherche.
|
| Les interactions entre plantes
(compétition, facilitation), et les interactions entre
les plantes et leur environnement physique sont toujours locales.
De ce fait, on peut prédire qu'il existe un réseau
complexe d'interactions entre la démographie d'une population
de plantes, la répartition spatiale des individus qui
la composent, et l'hétérogénéité
préexistante de l'écosystème. Le but de
ma thèse, réalisée au sein du laboratoire
"Fonctionnement et évolution des systèmes
écologiques" (Paris VI, ENS, CNRS), sous la direction
de J.-C. Menaut et Jacques Gignoux , a été de
mettre en évidence ce type d'interactions et d'en démêler
les mécanismes. Dans ce cadre, j'ai travaillé
sur le palmier Rônier (Borassus aethiopum) dans la savane
de Lamto en
Côte d'Ivoire. |
 |
|
 |

L'équipe
de techniciens de Lamto
|
|
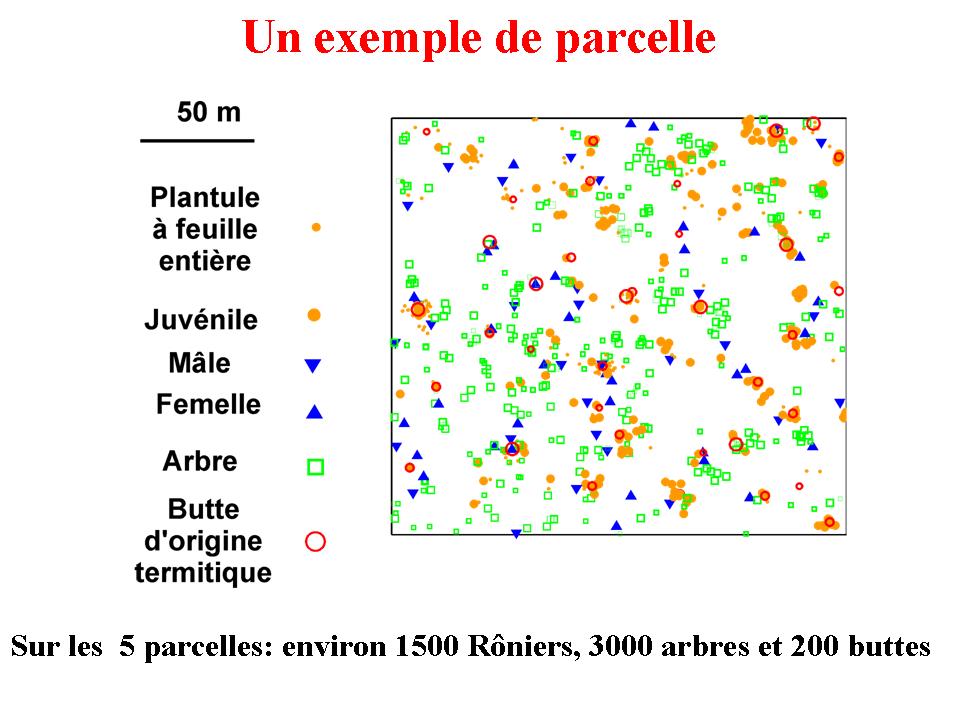
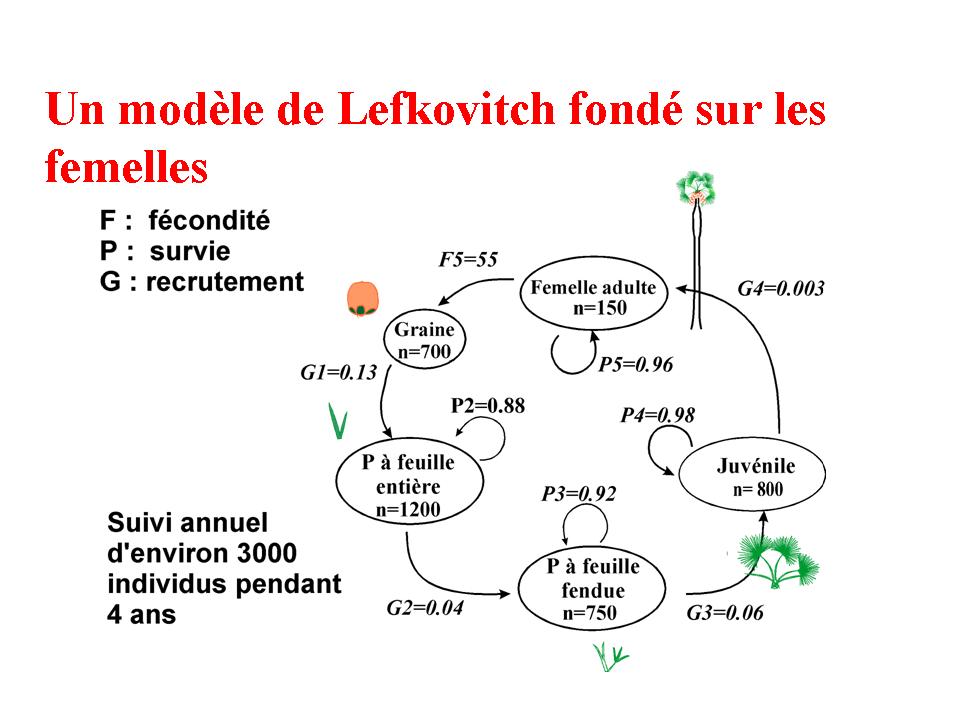
Dans un premier temps, le cycle de vie du Rônier
a été décrit, et modélisé à
l'aide d'un modèle matriciel (Barot
et al. 2000 JTE). Dans un second temps, une analyse exhaustive
de la répartition spatiale de la population de Rôniers
a été entreprise à l'aide des méthodes
de Ripley et de Diggle. Cette analyse a permis de construire un
scénario de démographie spatialisée pour le
Rônier, qui met en évidence les interactions locales
entre les palmiers et les taches de sol riche en nutriments (buttes
d'origine termitique et bosquets d'arbres), ainsi que les interactions
entre palmiers (Barot
et al. 1999 Ecology). Dans un troisième temps, ce scénario
a été testé à l'aide de données
démographiques tenant compte de la localisation des individus.
|
|
| D'une manière générale, la survie
et le recrutement des individus d'une population végétale
semblent conditionnés par de petites différences dans
la qualité de leurs voisinages qui conduisent à de petites
différences de croissance qui s'amplifient au fil des années.
Certains paramètres démographiques semblent ainsi déterminés
par la répartition spatiale des individus au stade considéré,
et donc en définitive par la répartition spatiale initiale
des graines (Barot
et al. 2003 JVS). |
| |
|
| Post-doctorat |
|

L'IIASA est hébérgé
dans un château du XVIIIème siècle
|
J'ai effectué un post-doctorat à l'International
Institute of applied Systems Analysis (Laxenburg, Autriche) au sein
de l'Adaptative dynamics Network. Mon travail a été
financé par la Communauté Européenne par l'intermédiaire
du réseau européen (European Training Network) ModLife
qui visait a réunir une masse critique de chercheurs pour
travailler, tant sur le plan théorique (modélisation)
qu'empirique sur l'évolution des histoire de vie. J'ai travaillé
sous la direction d'Ulf Dieckmann (chef du projet) et de Mikko Heino.
|
|

Gadus morhua |
 |
|
|
Mon projet de recherche personnel a porté
sur la dynamique des espèces de poissons de mer péchés
commercialement. Il est primordial d’étudier leur dynamique
et d’améliorer la qualité des prédictions
en résultant pour élaborer des stratégies de
pêche optimales : permettant à la fois de pêcher
un maximum de poissons et la durabilité de l’exploitation.
Ce type d’étude est indispensable dans le contexte
de l’augmentation de la population mondiale, et donc de la
pression exercée sur les écosystèmes naturels,
et de l’érosion de la biodiversité qui en est
souvent la conséquence. Mon travail a consisté à
analyser des données récoltées par les pêcheries
de morues (Gadus morhua) canadiennes et américaines.
Ces populations de morues ont été surexploitées
ce qui a abouti à l’effondrement des populations au
début des années 90, et à l’interdiction
quasi totale de la pêche. Mon travail visait à permettre
de comprendre mieux pourquoi les modèles n’ont pas
permis de prédire l’effondrement de ces populations
de morues et donc de le prévenir en diminuant à temps
les quotas de pêche.
|
L’hypothèse principale est que des
facteurs n’ont pas été pris en compte, par ces
modèles : des facteurs extérieurs comme des modifications
climatiques (température de l’eau), et des facteurs
‘‘internes’’ à la dynamique des populations
de morue, comme les variations de fécondité en fonction
de la taille des femelles. Il s’agit alors de déterminer
le rôle respectif de ces 2 types de facteur. C’est d’une
manière générale une opération critique
quand il s’agit de faire des prédictions sur les changements
que connaîtront des populations animales ou végétales
dans le cadre du changement global. Mon travail de thèse
sur le palmier Rônier, et l’ensemble de mon projet de
recherche sur la dynamique de la végétation en savane
ont en fin de compte un but similaire: déterminer l’influence
respective du forçage physique, et des paramètres
biologiques et physiologiques sur la dynamique globale des populations
considérées.
|
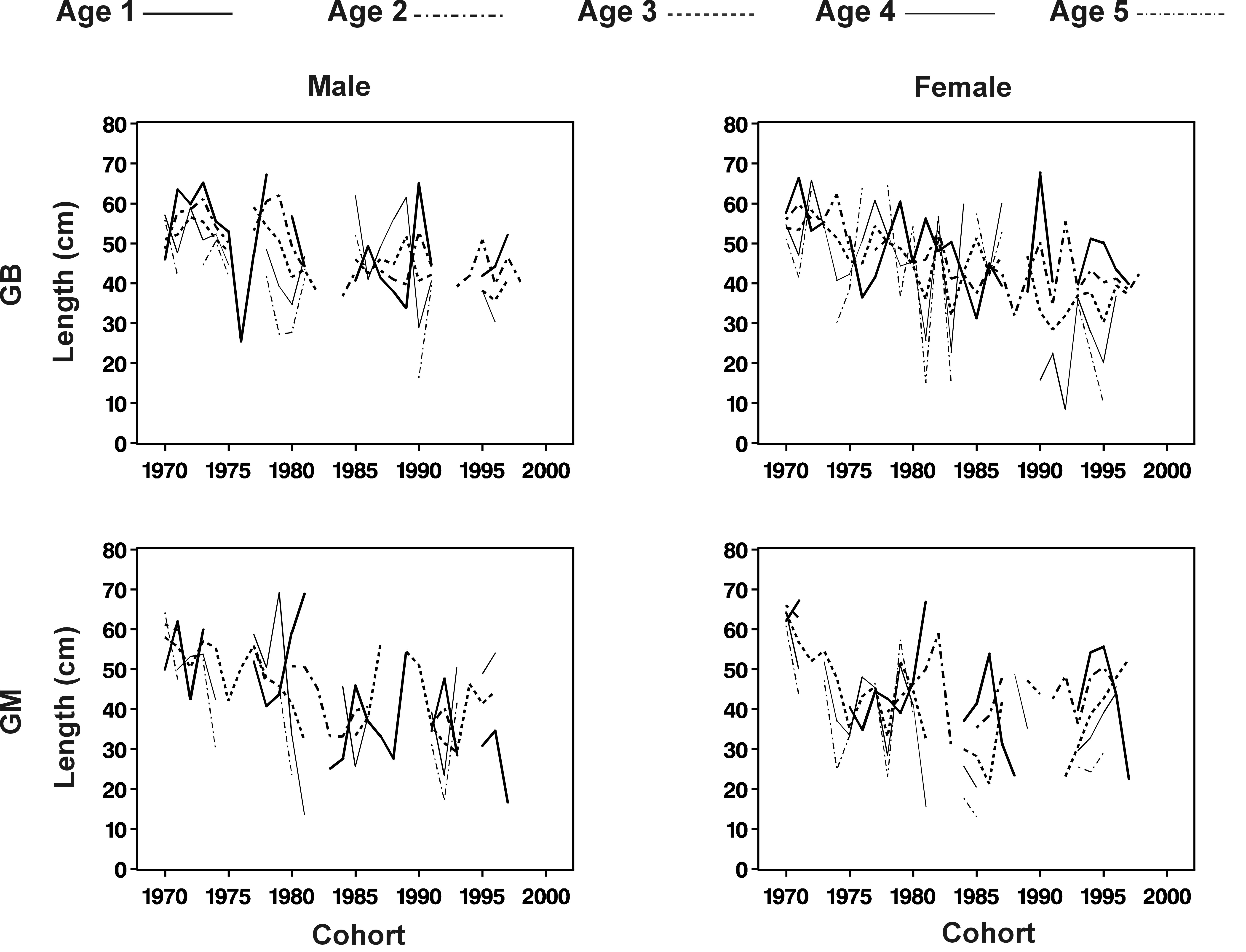 |
Normes de réaction
pour l'âge et la taille à la première
reprodcution. GB, Georges Bank; GM, Gulf of Maine. Estimations
séparées pour les mâles et les femelles
de 1970 à 1998. |
|
Plus spécifiquement, la pêche
est la principale source de mortalité pour les morues adultes
et juvéniles. La théorie de l’évolution
prédit que des forts taux de mortalité chez les juvéniles
devrait conduire à des modifications génétiques
menant à une reproduction de plus en plus précoce (‘‘de
façon à diminuer la chance de mourir avant de se reproduire’’).
C’est ce qui est apparemment observé chez la morue. Il
se pose alors un problème crucial: comment distinguer une diminution
évolutive de l’âge à la première
reproduction, d’une diminution uniquement due à la ‘‘plasticité
phénotypique’’ (capacité d’organismes
ayant les même gènes à présenter des phénotypes
différents selon les conditions physiques) et à des
modifications du milieu physique. Mon travail a ainsi consisté
à élaborer des méthodes statistiques permettant
une telle distinction à partir de longues séries temporelles
(cf. figure ci-dessus, distinction entre les 2 flèches déterminant
le phénotype)(Barot et
al 2004 EER). Cette distinction est primordiale sur le plan théorique
(mettre en évidence un phénomène évolutif),
mais aussi sur le plan pratique: des modifications génétiques
sont beaucoup plus difficilement réversibles que des modification
purement phénotypiques et il est important d’élaborer
des stratégies de pêche qui n’aboutiraient pas
d’une manière non réversible à pêcher
des poissons de plus en plus petits… Mon travail suggère
que la diminuation de l'âge et de la taille à la première
reproduction sont bien dus à un processus évolutif (Barot
et al 2004 Ecol. Apl.). Il est nécessaire d'effectuer le
même type d'étude sur d'autres populations pour conforter
ces résultats. J'ai obtenu des résultats semblables
pour le carrelet (Barot
et al. 2005 ICES J...). |
|
| |
| Modélisation
et faune du sol |
|
| L'écologie des sols utilise moins la modélisation
que les reste de l'écologie, comme on peut le voir en lisant
Pedobiologia, Soil Biology and Biochemistry, European Journal of Soil
Biology, ou Biology and Fertility of Soils. Pourtant la modélisation
est actuellement un outil indispensable pour tester de nouvelles hypothèses,
suggérer des mécanismes pour expliquer des patterns
particuliers, faire des prédictions quantitatives ou suggérer
de nouvelles expériences. A l'IRD, au sein de l'UMR 137, j'ai
développé des modèles au sujet des vers de terre.
Je suis parti dans deux directions complémentaires. (1) Modéliser
la dynamique spatiale des vers de terre pour comprendre pourquoi ils
ne sont pas répartis spatialement de manière homogène.
(2) Modéliser l'influence des vers de terre sur le recyclage
des nutrimens et voir comment cela peut influencer la production primaire.
|

Cadre de travail à Bondy |
|
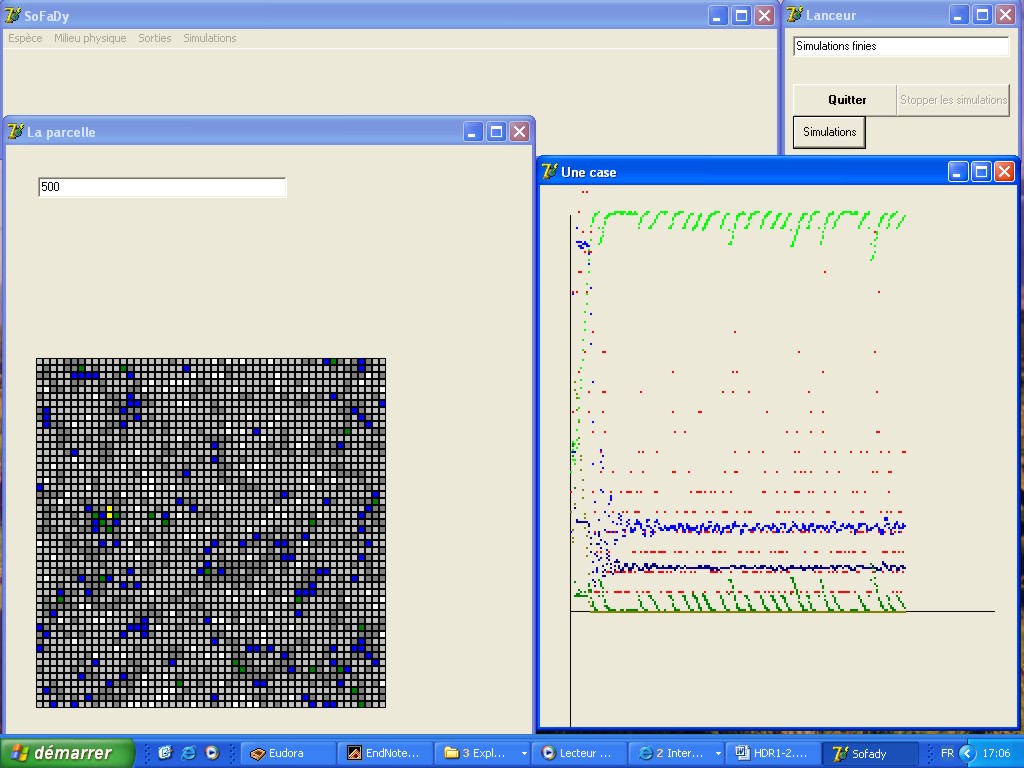
Interface du programme de simulation de
la démographie spatialisée des vers de terre |
La démographie et la répartition spatiale de la faune
du sol dépendent à la fois de facteurs démographiques
propres (comme la capacité de dispersion, la fécondité,
la survie) et de l’hétérogénéité
physico-chimique du sol. De plus, la faune du sol rétroagit
sur son environnement en modifiant le sol. Cela est en particulier
le cas pour les ingénieurs de l’écosystème
que sont les vers de terre (Jones et al. 1997 , Lavelle et al. 1997).
De part cette boucle de rétroaction, la dynamique d’une
population de vers de terre peut être extrêmement complexe.
En retour, cela rend difficile la prédiction des variations
temporelles et spatiales de l’état du sol (structure,
quantité et qualité de la matière organique)
sur lequel les vers agissent. Or de telles prédictions sont
indispensables pour comprendre certains processus écosystémiques
critiques dans le contexte du changement global : le stockage de carbone,
la minéralisation, ou l’évolution de la fertilité
sur le long terme. Potentiellement, une population de vers de terre
peut aider au maintien ou au rétablissement de la structure
du sol ou à l’inverse détruire cette structure
(Chauvel et al. 1999). |
| Avec Jean-Pierre Rossi j'ai construit un modèle
de simulation spatialement-explicite pour tester l'hypothèse
selon laquelle la démographie des vers de terre peut aboutir
à une répartition hétérogène (existence
de grandes zones d'environ 10 m de diamètre où la densité
en vers est beaucoup plus forte que leur densité moyenne) sans
forçage extérieur. L'idée est que des effets
de densité dépendance négative combinés
avec une faible mobilité peut produire de l'hétérogénéité.
Le modèle est une sorte d'automate cellulaire qui permet de
suivre dans le temps et dans chaque cellule la densité de vers
de terre et un paramètre du sol (Barot et al. 2007 SBB). Ce modèle a été
paramétré pour Millsonia anomala qui a été
étudié de manièreexhaustive par Patrick Lavelle
dans la savane de Lamto en Côte D'Ivoire. |
Sorties du programme de simulation de la
démographie spatialisée des vers de terre (cartographie
et variogramme) |
| |
|
| |
|
|
| Effets des vers de terre sur la croissance et la démographie des plantes |
|
J’ai obtenu un financement sur 3 ans (2006-2009) du programme ANR-Jeune chercheur (projet SolEcoEco). L’idée de ce projet est que l’écologie des sols manque d’un cadre conceptuel général permettant à la fois d’interpréter un corpus très riche d’observations empiriques et de suggérer de nouvelles expériences clefs. Je pense qu’un tel cadre devrait être fondé sur l’idée que les interactions entre plantes et organismes du sol (vers de terre, termites, bactéries …) ont été façonnées par des pressions de sélection et donc des processus évolutifs. Ce projet a donné lieu a trois thèses que j’ai encadrées : Kam-Rigne Laossi (tchadien, thèse soutenue en Janvier 2009) et de Diana Noguera (colombienne, partenariat avec le CIAT de Cali, soutenance en Septembre 2009) et Simon Boudsocq (voir la section modélisation, thèse soutenue en Janvier 2010).En 2009 et 2010 nous avons achevé les analyses, la rédaction d’articles et leur publication correspondant à ces trois thèses. |
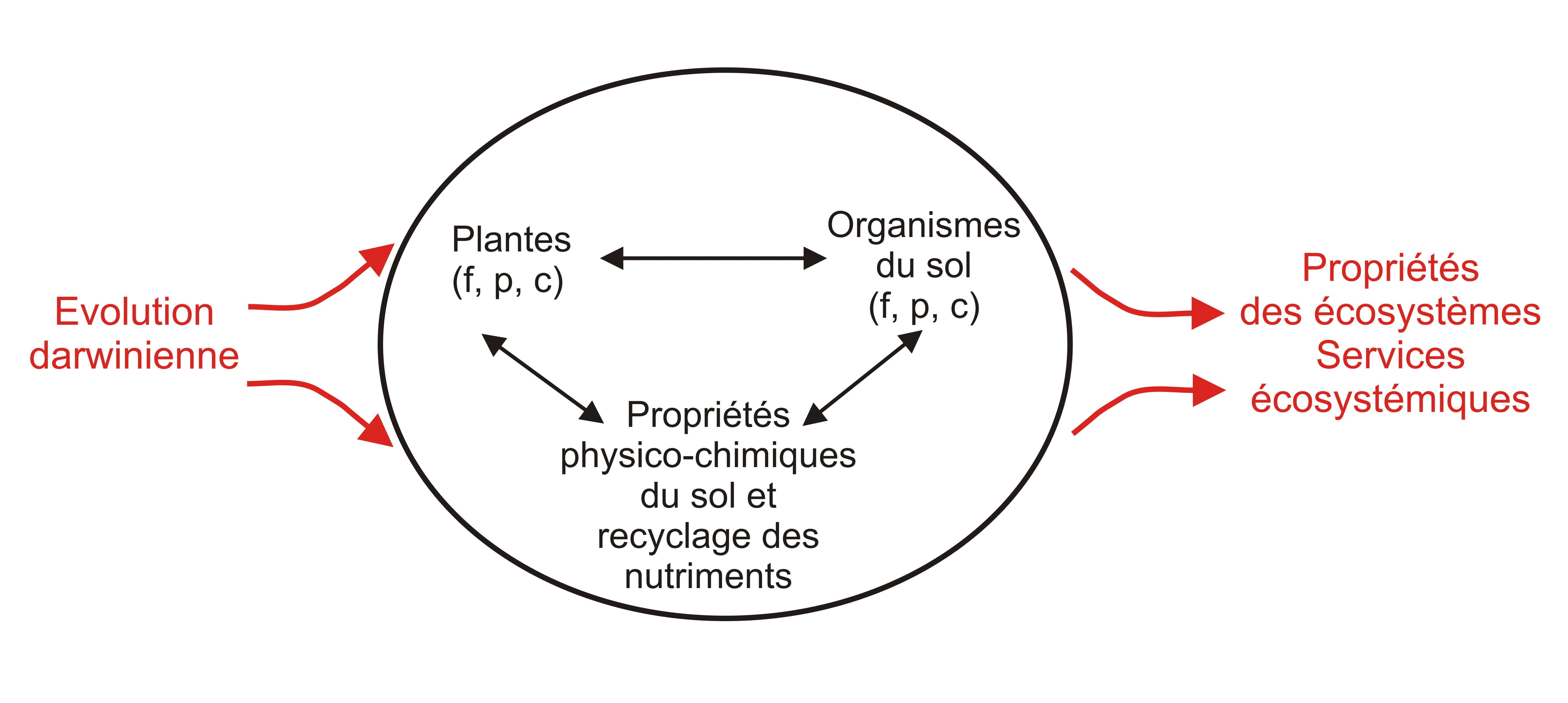
Schéma conceptuel général montrant comment les plantes, les organismes du sol et le sol interagissent, comment ces interactions sont soumises à l'évolution, ce qui détermine en partie les propriétés des écosystèmes |
|
 Mésocosmes contenant une communauté de quatre espèces de plantes annuelles Mésocosmes contenant une communauté de quatre espèces de plantes annuelles |
Les vers de terre constituent une composante majeure de la faune du sol et il est désormais bien établi qu’ils affectent la croissance des plantes. Bien que les espèces de plantes répondent différemment aux vers de terre au cours d’un cycle de vie, très peu de travaux ont étudié l’effet des vers de terre sur la compétition entre plantes et sur leur démographie. Les vers de terre peuvent affecter la stratégie d’allocation des ressources des plantes et ainsi, influencer leur survie et leur fécondité. Ils pourraient donc affecter la démographie de différentes espèces de plantes et donc, au-delà d’une génération, la structure des communautés végétales. K.-R. Laossi a étudié dans une série d’expériences l’effet d’un ver endogé (A. caliginosa) et d’un ver anécique (L. terrestris) et de leur interaction sur quatre plantes annuelles (T. dubium, V. persica, P. annua et C. glomeratum) du niveau individuel (réponse de chaque espèce végétale considérée individuellement) au niveau de la communauté végétale (réponse des plantes en situation de compétition au sein d’une communauté végétale). Nous avons ainsi montré que les vers de terre affectent la croissance des plantes et que cet effet varie en fonction de l’espèce végétale et du type de sol utilisé. De plus, ils modulent la compétition entre plantes sur le court (accululation de biomasse) et le long terme (fécondité et germination des graines) (Laossi et al. 2009 SBB) et leurs effets sur les plantes peuvent être transmis à la génération suivante, par un effet maternel (Laossi et al. 2010 SBB). Enfin, ils modifient la démographie des plantes en influençant leur fécondité, leur survie et la germination des graines. Dans une expérience en mésocosmes qui a maintenu pendant trois générations des communautés des quatre plantes annuelles avec et sans vers, nous avons pu vérifier que les mécanismes démographiques mis en évidence dans les expériences à court terme modifient bien la structure de la communauté sur le long terme (Laossi et al. 2011 Pedobiol.). |
 Aporrectodea caliginosa Aporrectodea caliginosa |
|
La fertilité des sols tropicaux est souvent peu durable dans les systèmes de cultures et les pâturages. Ce problème de fertilité est en grande partie du à la très rapide décomposition de la matière organique dans ces sols et à leur faible capacité à retenir les nutriments minéraux. De plus, dans un contexte où une grande partie des agriculteurs ont peu accès aux intrants (engrais minéraux) il est important de trouver des méthodes de gestion durable et nécessitant peu d’intrants. Une solution pourrait être d’enrichir le sol en charbon de bois fragmenté pour créer des ‘‘terres noires’’ comme celles découvertes récemment en Amazonie et qui auraient été implantées il y a des siècles par des groupes amérindiens aujourd’hui disparus. Une autre solution pourrait être de promouvoir des systèmes agricoles favorables aux vers de terre dont les activités augmentent généralement la croissance des plantes. Les mécanismes impliqués commencent à être connus tant pour les vers que pour les terres noires mais les interactions entre vers et terres noires n’ont jamais été abordées. D. Noguera a étudié dans une série d’expériences l’effet de vers de terre (P. corethrurus), du charbon bois et de leur interaction sur le croissance du riz: 1) réponse du riz dans différents types du sol (expérience en microcosmes), 2) réponse des 5 variétés du riz dans un type sol et dans deux niveaux de fertilisation minérale (expérience en microcosmes), 3) réponse physiologique et cellulaire du riz à la présence de vers de terre et de charbon bois et 4) effet des vers de terre sur la formation de terre noire et leur fertilité (expérience en mésocosmes). Nous avons ainsi montré que les vers de terre et le charbon bois affectent la croissance des plantes et que les effets vers de terre et charbon bois varient en fonction du type de sol utilisé et diminuent avec la fertilisation minérale (Noguera et al. 2010 SBB).
|
 Microcosmes contenant chacun un pied de riz Microcosmes contenant chacun un pied de riz |
|
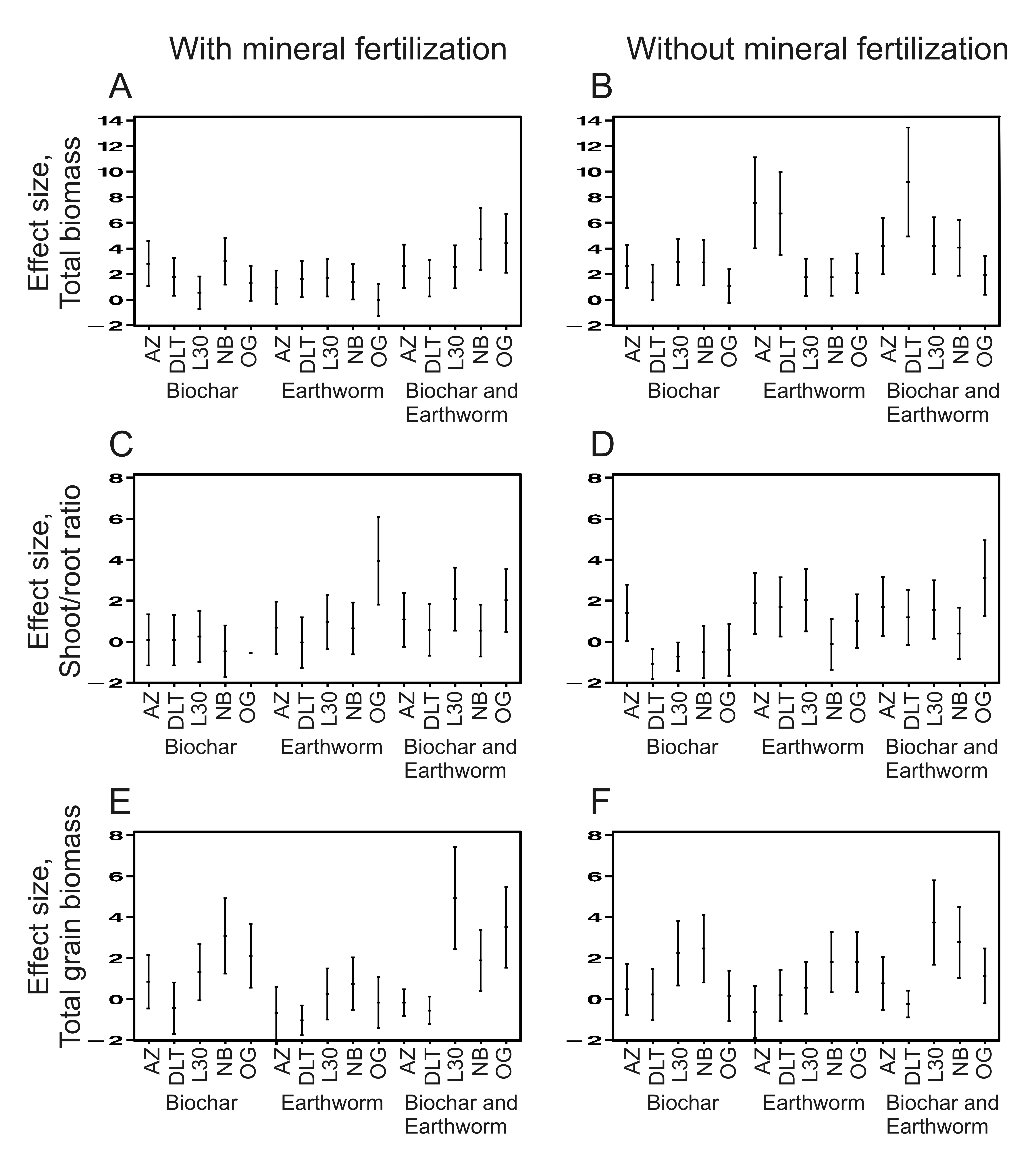 Taille des effet des différents traitements observés pour les différentes variétés de riz Taille des effet des différents traitements observés pour les différentes variétés de riz |
Les résultats d’une deuxième expérience montrent que les différentes variétés de riz répondent différemment à la présence de charbon bois et de vers. Cette expérience a confirmé nos hypothèses. Les cinq variétés de riz ont bien des réponses très différentes aux vers, au charbon de bois et à la combinaison des deux. L’accroissement moyen de la production de grains est d’environ 90% et 30 % respectivement, avec et sans fertilisation minérale, pour l’ensemble des traitements et des variétés. Ce bénéfice lié aux vers de terre ou au charbon de bois est multiplié par quatre (et plus) quand on choisit la meilleure combinaison de traitement et de variétés (gain de 430 % et de 350% respectivement avec et sans fertilisation). De plus, (1) la variété de riz la plus rustiques, sélectionnées localement en Colombie par les agriculteurs, est la variété qui répond le mieux, en termes de biomasse totale, aux vers de terre ; (2) la variété répondant le mieux en terme de production de grain est une variété moderne sélectionnée pour l’agriculture intensive. Ces résultats montrent que l’agro-écologie bénéficierait grandement de la sélection de variétés spécifiques. D’une manière générale, ils suggèrent aussi que la sélection variétale moderne à pu faire perdre aux plantes cultivées la capacité d’interagir finement et positivement avec les organismes du sol. A l’inverse, sélectionner de nouvelles variétés pour leur capacité à bénéficier de ces organismes et de l’hétérogénéité du sol pourrait reconstituer les boucles de rétroaction positive constituées au cours de l’évolution, dans les écosystèmes naturelles, entre les plantes et organismes du sol. Cela pourrait augmenter la durabilité de l’agriculture. Enfin, d’un point de vu plus fondamental, nos résultats suggèrent que les vers de terre peuvent constituer une pression de sélection significative pour les plantes, même si les caractéristiques sélectionnées ne sont pas encore connues (architecture et dynamique racinaire particulière ?)(Noguera et al. 2011 Ecol. Appl.). |
|
| |
|
|
| Recyclage des nutriments, modélisation et évolution |
|
| La question de l’importance du recyclage des nutriments minéraux est une très vieille question en écologie. D’une certaine manière elle est à la base de l’écologie des écosystèmes et de l’idée que les écosystèmes constituent des systèmes dont tous les acteurs sont liés. Ainsi, les plantes absorbent des nutriments minéraux (l’azote par exemple), la matière organique issue des feuilles et des racines mortes retourne au sol, mais les nutriments qu’elle contient sont essentiellement rendus disponibles pour les plantes après minéralisation, sous l’influence de la microflore des sols, mais aussi de la méso- (par exemple les enchytréides) et macrofaune (par exemple les vers de terre) des sols qui participent à la décomposition de la matière organique. On reconnaît cependant de plus en plus que les systèmes écologiques sont des systèmes ouverts et que la dynamique d’un écosystème dépend aussi des entrées (fixation de l’azote atmosphérique, dépôt de nutriments apportés par le vent et les pluies) et des sorties de nutriments (lessivage, érosion). On peut alors se poser une question théorique pour laquelle aucune réponse tranchée n’a encore été apportée. Quel est le facteur le plus important pour la production végétale, et donc d’une certaine manière pour l’efficacité d’un écosystème ? L’efficacité du recyclage, c'est-à-dire la capacité d’un écosystème à conserver ses nutriments en en limitant les pertes et éventuellement à capter des nutriments venant de l’extérieur ? Ou la rapidité du recyclage, c'est-à-dire le temps que mettent les nutriments à parcourir une boucle complète de recyclage ? Ces questions ne sont pas purement théoriques, elles sont liées à des applications très importantes. Par exemple, l’agriculture moderne a surtout misé sur l’augmentation des entrées de nutriments (engrais), en se préoccupant relativement peu des sorties. On estime que les agroécosystèmes consomment 77 millions de tonnes d’azote minéral par an mais que le lessivage de sols cultivés conduit à 88 millions de tonnes de perte, et incidemment à la pollution de nombreuses nappes phréatiques et à l’eutrophisation de nombreux lacs et courts d’eau. Dans ce cadre je me suis intéressé à deux questions plus élémentaires. Quel est l’effet de la macrofaune du sol, et en particulier des vers de terre sur le recyclage des nutriments, sa rapidité et son efficacité ? Les plantes peuvent-elles avoir intérêt à inhiber la nitrification? |
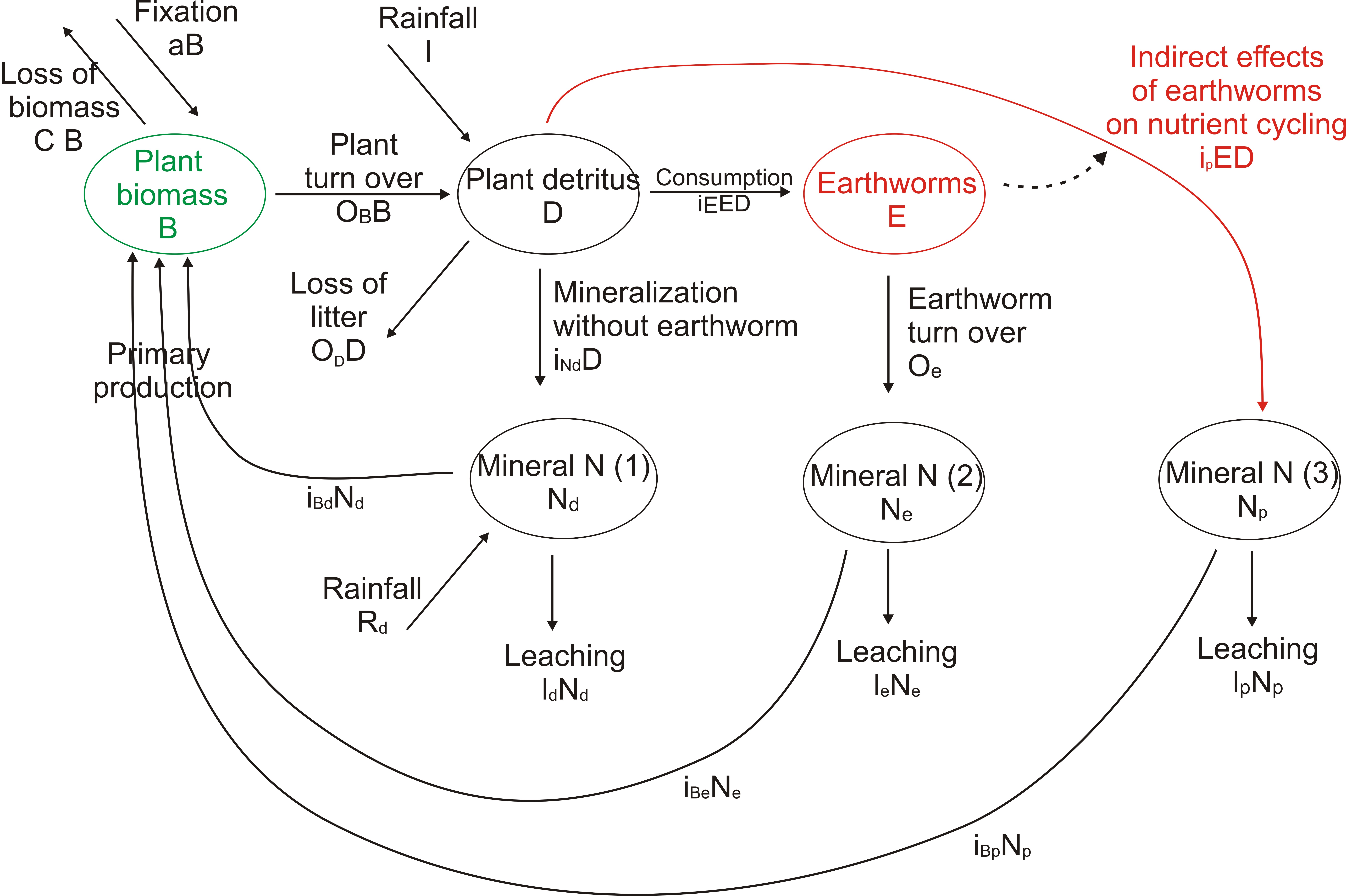 Shéma décrivant notre modèle de recylage des nutriments par les vers de terre Shéma décrivant notre modèle de recylage des nutriments par les vers de terre |
|
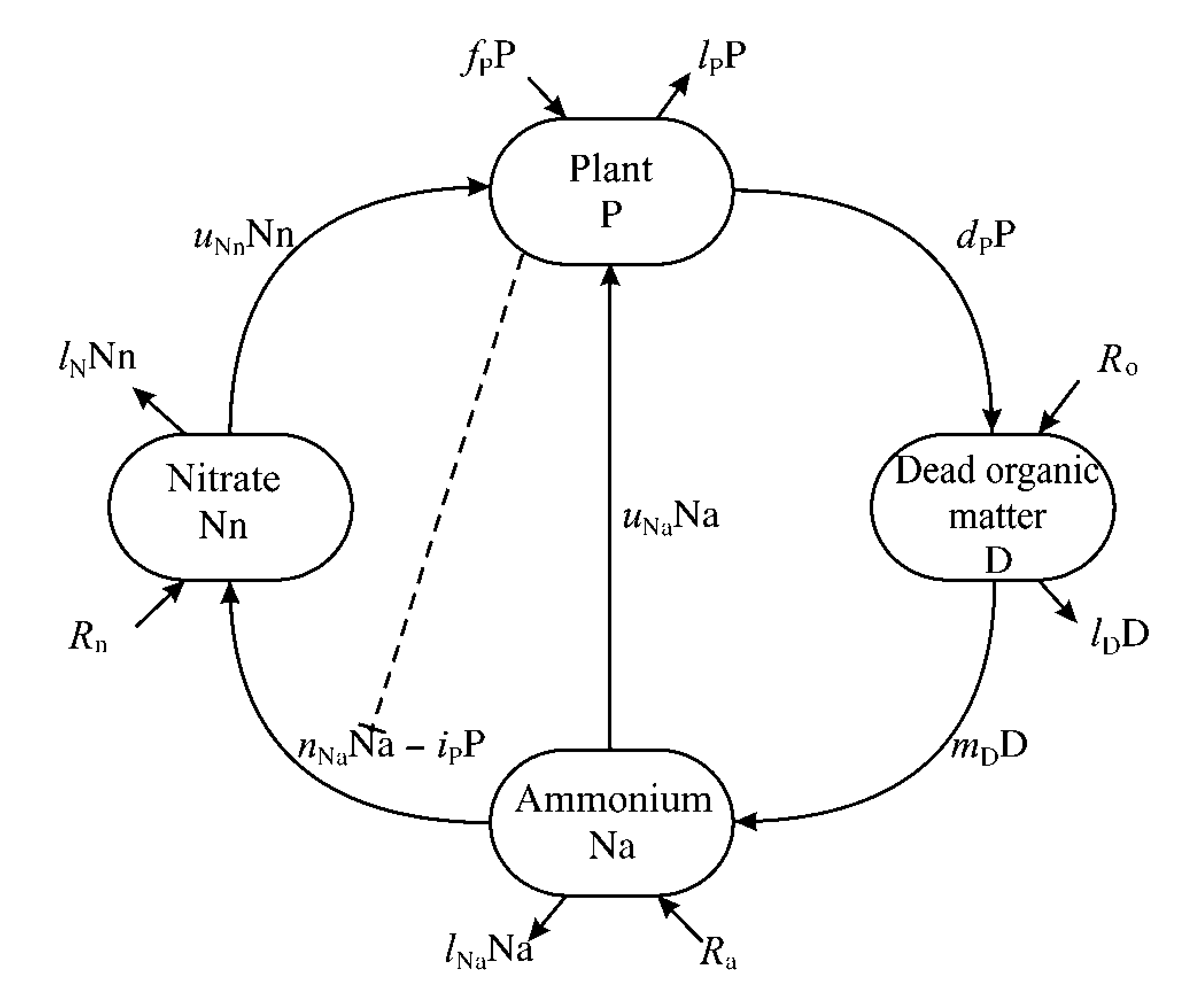 Shéma décrivant notre modèle de recylage de l'azote et de l'inhinition de la nitrification Shéma décrivant notre modèle de recylage de l'azote et de l'inhinition de la nitrification |
Le principal résultat est que les vers améliorent la croissance des plantes, sur le long terme, si et seulement si ils améliorent l’efficacité du recyclage des nutriments, c'est-à-dire si ils diminuent les pertes de nutriments pour l’écosystème et en augmentent les gains. Un second résultat, moins attendu, montre que si les vers augmentent l’efficacité du recyclage ils augmentent aussi leur propre biomasse, et qu’à chaque fois que les vers augmentent leur biomasse en augmentant l’efficacité du recyclage ils augmentent aussi la croissance des plantes. Cela est dû d’une part au fait que quand l’efficacité du recyclage augmente, une fois l’équilibre atteint (pour chaque compartiment du modèle les entrées de nutriments sont égales aux sorties), la quantité total de nutriments présents dans l’écosystème augmente nécessairement. Cela est dû d’autre part au fait que tous les nutriments passent par le ‘‘compartiment plante’’ au cours de leur recyclage, mais que seule une partie de ces nutriments passent par le ‘‘compartiments vers de terre’’. Ces derniers résultats sont intéressants dans la mesure où ils suggèrent que si certaines activités d’ingénieurs des écosystèmes des vers ont été sélectionnées au cours de l’évolution parce qu’elles augmentent la biomasse de vers de terre, ces activités augmentent aussi la croissance des plantes. Cela suggère ainsi que quand les vers de terre augmentent la croissance des plantes cela n’est pas forcément dû a des effets secondaires de leur mode de vie, mais que les mécanismes favorisant la croissance des plantes ont pu être sélectionnés parce qu’ils sont aussi favorables aux vers. Cela démontre la possibilité d’une coévolution entre vers et plantes qui aurait abouti à une forme de mutualisme. |
|
| Un second modèle (Boudsocq et al. 2009) a été développé par Simon Boudsocq au cours de sa thèse (soutenance en décembre 2009). De nombreuses graminées pérennes africaines inhibent la nitrification. C’est le cas d’Hyparrrhenia diplandra qui a été très étudiée dans la savane de Lamto en Côte d’Ivoire. Le modèle a en premier lieu permis de déterminer la condition générale sous laquelle réduire la nitrification augmente la production primaire : en fait il s’agit que le nitrate soit soumis à plus de perte que l’ammonium (par dénitrification et par lixiviation). Dans ce cas, inhiber la nitrification limite les pertes d’azote pour l’écosystème. Cette condition est très largement vérifiée. Le modèle a aussi permis de montré, en le paramétrant pour la savane de Lamto, que l’inhibition de la nitrification était l’hypothèse la plus vraisemblable pour expliquer la forte productivité primaire du couvert herbacé à Lamto. Ces résultats renforcent l’intérêt déjà existant pour le contrôle de la nitrification en agriculture. Nous travaillons maintenant sur les conditions sous lesquelles l’inhibition de la nitrification peut être sélectionnée au cours de l’évolution (au sens darwinien). Par ailleurs Simon a aussi développé un modème montrant que, logiquement, le préférence d'une plante pour l'ammonium ou le nitrate interagit avec sa capacité à inhiber la nitrification (Boudsocq et al. 2012). C'est un résultat original dans la mesure où les plantes peuvent consommer l'azote minéral sous ces deux formes mais que, par défaut, on supose souvent que très majoritairement du nitrate est absorbé. En fait, une plante obtient toujours son optimum de biomasse en consommant à la fois du nitrate et de l'ammonium. C'est ce qui permet de limiter au maximum les pertes d'azote minéral et permet toujours d'avoir accès à une ressource en azote importante vu que la minéralisation de la matière organique donne d'baord de l'ammonium. Dans cet article, nous montront aussi que cette préférence pour l'ammonium ou le nitrate influence aussi la capcité d'une espèce de plante à envahir une population d'une autre espèce ou à résister à une invasion. Cela peut contrinuerà expliquer, par exemple, l'invasion de certaines savanes australiennes par des graminées africaines |
|
|
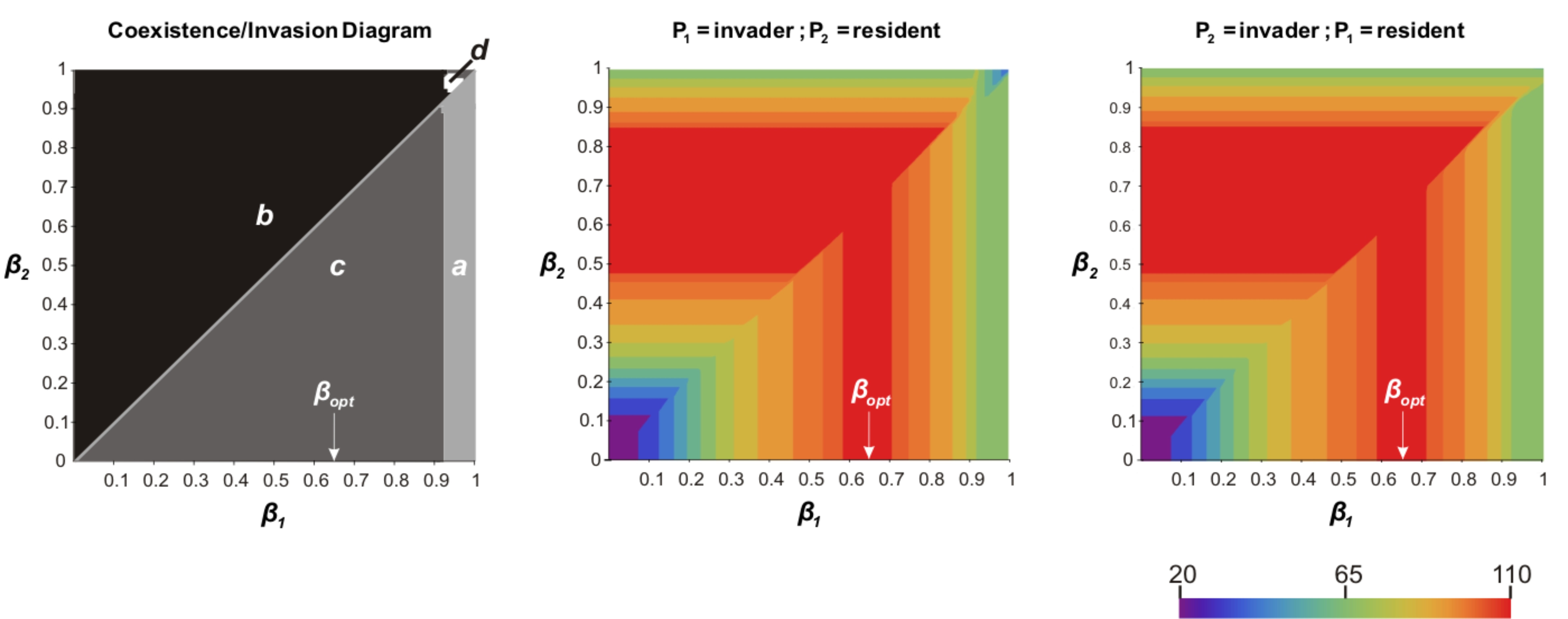 |
Invasion réciproque entre deux plantes (zones plus oumoins sombres) différant par leur préférence pour l'ammonium (beta) et biomasses en résultant (gradient de couleur) |
|
| Simon a développé un autre modèle (Boudoscq et al. 20010) visant à analyser l'évolution de l'investiment des plantes pour l'absorption des nutriments minéraux. Pour cela, il a utilisé les techniques mathématiques développées par la dynamique adaptative.L'idée générale est d'étudier comment l'évolution peut façonner certaines propriétés des écosystèmes. En faisant l'hypothèse que les plantes doivent faire un compromis entre la vitesse d'absorption des nutriments et le temps de résidence des nutriments dans leur biomasse, nous avons montré que trois issues évolutives étaient possibles: (1) un équilibre évolutif est atteint, (2) la biomasse de la plante diminue continuellement ce qui aboutit à un suicide évolutif, (3) au cours de l'évolution le recyclage des nutriments devient tellement efficace que le nutriment inititalemen limitant s'accumule dans l'écosystème et devient non-limitant. A l'équilibre évolutif, la stratégie développée au cours de l'évolution minimise le stock de nutriments minéraux (supportant à l'échelle évolutive le R* de Tillman) mais cette stratgie ne maximise pas la biomasse de plante ou la production primaire. |
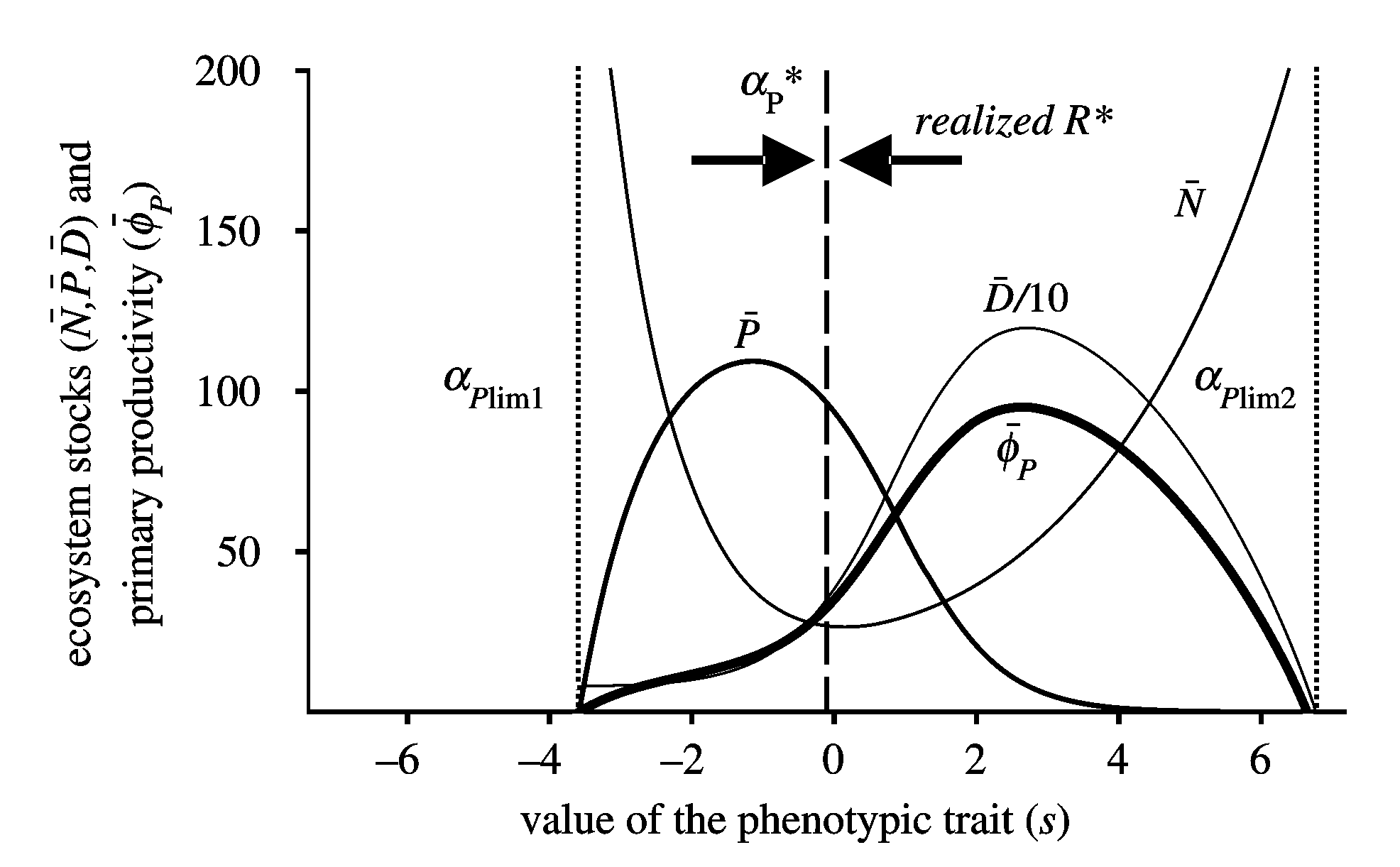 Effet de l'investissement d'une plante pour la capture des nutriments sur les différents compartiments écologiques. Dans ce cas l'évolution conduit à un équilivre évolutif qui minimise N mais ne maximise pas P Effet de l'investissement d'une plante pour la capture des nutriments sur les différents compartiments écologiques. Dans ce cas l'évolution conduit à un équilivre évolutif qui minimise N mais ne maximise pas P |
|
| |